


Abstract
The mechanism of the hypocholesterolemic action of S-8921, methyl 1-(3,4-dimethoxyphenyl)-3-(3-ethylvaleryl)-4-hydroxy- 6,7,8-trimethoxy-2-naphthoate, was examined in rats. In diet-induced hypercholesterolemic rats, 2 weeks oral administration of S-8921 dose- and time-dependently decreased plasma cholesterol level in the daily dose range of 0.1to10 mg/kg. Results with the dual-isotope plasma ratio method indicated that S-8921 inhibits cholesterol absorption from the intestine and enhances its elimination from the body. The in situ loop method showed that S-8921 does not inhibit the absorption of cholesterol from rat jejunum, clearly inhibits active absorption of taurocholic acid (TCA) and glycocholic acid (GCA) from rat ileum and does not inhibit passive absorption of cholic acid (CA) from the rat jejunum. In rat ileal brush-border membrane vesicles, S-8921 inhibited the sodium-dependent uptake of TCA in a concentration-dependent manner with IC50 of 2.1 μM, not the Na+-dependent d-glucose andl-alanine uptake. These results suggest that S-8921 is a potent, selective inhibitor of the Na+-dependent bile acid transport system in the ileal mucosal cell brush-border membrane, and this inhibition is the mechanism by which this drug decreases intestinal bile acid reabsorption to result in a significant decrease of plasma cholesterol.
Hypercholesterolemia, especially a high level of LDL cholesterol, is a major risk factor for atherosclerosis which contributes to coronary heart disease. Randomized clinical trials have demonstrated that intensive reduction of plasma LDL cholesterol levels could halt and reverse atherosclerosis and decrease the incidence of coronary heart disease (Lipid Research Clinics Program, 1984a, b; Blankenhorn et al., 1987; Brownet al., 1990). Hence, many types of cholesterol-lowering drugs have been developed to ameliorate hypercholesterolemia to prevent the incidence and progression of atherosclerosis. Combination therapy with agents with different hypocholesterolemic mechanisms such as bile acid sequestrants and inhibitors of the HMG CoA reductase has achieved greater synergistic reduction in blood cholesterol level (Vega and Grundy, 1987; Leren et al., 1988). Therefore, much effort is still being directed to finding agents with other hypocholesterolemic mechanisms.
Recently, we found that S-8921, a lignan derivative, has a potent cholesterol-lowering activity in vivo on screening with mice and an antioxidative property against LDL oxidation in vitro. Another lignan derivative was previously reported to reduce serum cholesterol in rats fed diet containing cholesterol and cholate (Takashima et al., 1994). However, the precise mechanism of this agent is not clear, although its hypocholesterolemic activity is suggested to be associated with cholesterol and/or bile acid absorption in the intestine. The purpose of the present study was to elucidate the mechanism of the hypocholesterolemic action of S-8921 in rats. The present experiments examined the effects of S-8921 on plasma cholesterol concentrations in vivo, on cholesterol absorption and elimination by the dual-isotope plasma ratio method (Zilversmit, 1972; Zilversmit and Hughes, 1974; Samuel et al., 1978), on bile acid transport by rat ileal brush-border membrane vesicles in vitro and rat ileal loop in situ and on bile acid excretion in vivo.
Materials and Methods
Materials.
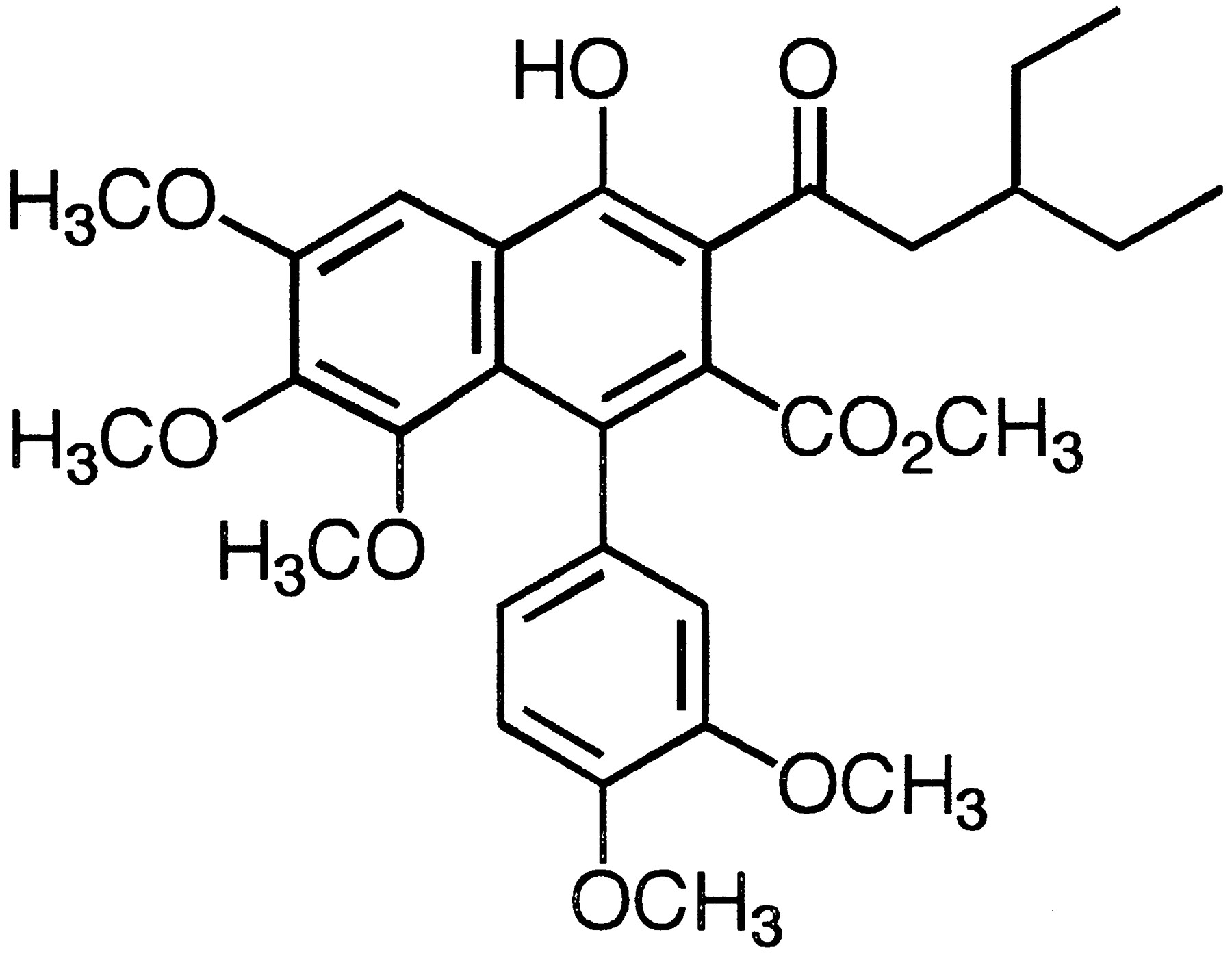
S-8921 [methyl 1-(3,4-dimethoxyphenyl)-3-(3-ethylvaleryl)-4- hydroxy- 6,7,8-trimethoxy-2-naphthoate] was synthesized at Shionogi Research Laboratories, Osaka, Japan. The chemical structure of this compound is shown in figure 1. [4-14C]Cholesterol (1.85 GBq/mmol, 99.1%), [1a,2a(n)-3H]cholesterol (1.78 TBq/mmol, 99.1%), tauro[carbonyl-14C]CA, sodium salt (1.96 GBq/mmol, 95.1%), [1-14C]GCA, sodium salt (2.07 GBq/mmol, 98.7%),d-[6-14C]glucose (2.01 GBq/mmol, 99.7%) and l-[U-14C]alanine (5.9 GBq/mmol, 99.3%) were purchased from Amersham International plc (Buckinghamshire, England). [Carboxyl-14C]CA (1.8 GBq/mmol, 99%) was from NEN Research Products (Wilmington, DE). The radiochemical purities of the compounds were confirmed by TLC. HEPES, Tris, TCA, sodium salt, monoolein, polysorbate 80 and gum arabic were from Nacalai Tesque (Kyoto, Japan). Sesame oil was from Maruishi Pharmaceutical Co., Ltd. (Osaka, Japan) and other chemicals were of analytical or reagent grade.

Chemical structure of S-8921.
Animals.
Male Sprague-Dawley rats (10–11 weeks old) were purchased from Clea Japan, Inc. (Tokyo, Japan). The animals were housed in a room with controlled light (on from 8 a.m. to 8p.m.), temperature (23 ± 1°C) and humidity (55 ± 10%) and maintained on commercial chow (CA-1 pellets, Clea Japan) and water ad libitum.
Hypocholesterolemic effect of S-8921.
Rats were fed CA-1 chow supplemented with 1% cholesterol (Nacalai Tesque, Inc,. Kyoto, Japan) and 0.5% sodium CA (Nacalai Tesque, Inc., Kyoto, Japan) for 7 days, and then were divided into groups having the same average plasma cholesterol level. Together with continuous feeding of cholesterol-CA, S-8921 was administered as a suspension in 5% gum arabic (2 ml/kg) orally for 14 days at 9 a.m., except on the day of blood collection when S-8921 was given after blood sampling. Control animals were administered only 5% gum arabic as a vehicle. Blood was collected from the orbital vein under light ether anesthesia at 1p.m. from rats fasted for 4 hr before blood sampling. Plasma total cholesterol and HDL cholesterol were measured by an enzymatic method with commercial assay kits, Determiner TC555 (Kyowa Medex, Tokyo, Japan) and HDL cholesterol test Wako (Wako Pure Chemical Industries, Ltd., Osaka, Japan), respectively. Plasma VLDL + LDL cholesterol was calculated by subtracting HDL cholesterol determined as heparin-manganese-precipitable lipoprotein cholesterol from total cholesterol. A group of nontreated animals was kept to determine the base-line level of plasma cholesterol in ordinary chow.
Effect of S-8921 on cholesterol absorption and elimination (dual-isotope plasma ratio method).
Chylomicrons labeled with [14C]cholesterol were obtained from the thoracic duct lymph of anesthetized rats intraluminally administered 1.3 ml of mixed micellar solution containing [14C]cholesterol (1.48 MBq, 2 mM), Na TCA (10 mM), monoolein (5 mM) and lysolecithin (3.3 mM). The lymph was defibrinated. The oral dose of [3H]cholesterol (1.85 MBq) was prepared by dissolving 44 mg of cholesterol in 715 mg of sesame oil. The oil phase was then emulsified by sonication in 12 ml of Na TCA (5 mM) aqueous solution. S-8921 was administered as a suspension in 5% gum arabic (2 ml/kg) orally to rats once a day for 9 successive days at a dose of 1 mg/kg or 10 mg/kg. On the third day, 2 hr after administration of S-8921, a [3H]cholesterol aqueous emulsion (12 × 106 dpm, 4.7 mg/kg) was given orally to rats, then [14C]cholesterol-labeled chylomicrons (5 × 106 dpm/kg) were injected immediately into the tail vein. At a selected time after administration, the animals were anesthetized lightly with ethyl ether and blood was obtained from the tail vein in heparinized syringes. Blood samples were centrifuged and the plasma was separated.
Effect of S-8921 on cholesterol absorption (in situloop method).
The rats were anesthetized with ethyl urethane (1.4 g/kg s.c.), and thoracic ducts were cannulated by a modification of the method of Bollman et al. (1948), with use of a 20-cm length of polyethylene tubing (PE50, Becton Dickinson, Parsippany, NJ). Closed loops of jejunum (15 cm long from Treitz toward the distal site) were prepared by ligation at both ends. One milliliter of S-8921 aqueous suspension (10 μmol) was administered into the loop. The animals were kept on a warm plate at 38°C. After 2 hr, 1 ml (2 μmol as the dose of cholesterol) of mixed micellar solution containing [14C]cholesterol (2 mM), Na TCA (10 mM), monoolein (5 mM) and lysolecithin (3.3 mM) was instilled into the same loop, and the lymph was collected during 6 hr. Three milliliters of saline solution was injected into the tail vein at 1, 3 and 5 hr after dosing as a supply of water.
Effect of S-8921 on bile acid excretion (in vivo).
S-8921 aqueous suspension was administered orally to rats once a day for 3 successive days at a dose of 10 mg/kg. On the third day, 2 hr after administration of S-8921, [14C]TCA saline solution (5 μmol) was given orally to rats, and the feces were collected for 2 days.
Effect of S-8921 on bile acid absorption (in situloop method).
The rats were anesthetized with ethyl urethane and the tail vein was cannulated with a PE-50 catheter. A closed loop (7–8 cm) of the jejunum or the terminal ileum was made by ligating both ends. S-8921 aqueous suspension (0.5 μmol) was administered into the loop. In the comparable experiment, S-8921 (0.5 μmol) dissolved in saline containing 10% polysorbate 80 was injected into the tail vein of rats. The animals were kept on a warm plate at 38°C. After 1 hr, a second cannula filled with dilute heparin was introduced into the mesenteric vein governing the test segment. Immediately, 0.5 ml of 10 mM [14C]bile acid saline solution (5 μmol as the dose of bile acid) was instilled into the same loop, and the mesenteric blood was collected in the test tube for 30 min after dosing, with blood transfusion being given via the tail vein.
Effect of S-8921 on bile acid transport (in vitro).
Brush-border membrane vesicles were prepared from rat small intestine by the method of Beesley and Faust (1979). Jejunal and ileal brush-border membrane vesicles were obtained from the most proximal and most distal 20 cm of the rat small intestine, respectively. Brush-border membrane vesicles were freshly prepared on the day of use and suspended in 400 mM mannitol and 20 mM HEPES-Tris (pH 7.5) with the protein concentration at 2 mg/ml. The vesicle was preincubated for 1 min at 25°C. TCA uptake was initiated at zero time by addition of 70 μl of substrate medium containing 100 mM mannitol, 150 mM NaCl, 20 mM HEPES-Tris (pH7.5) and 50 μM [14C]TCA to 50 μl of vesicles at 25°C.d-Glucose or l-alanine uptake was initiated at zero time by the addition of 70 μl of substrate medium containing 100 mM mannitol, 0.1 mM MgSO4, 100 mM NaSCN, 1 mM HEPES-Tris (pH 7.5) and 22.9 μMd-[14C]glucose or 0.008 μMl-[14C]alanine to 50 μl of vesicles at 25°C. After various time intervals, the reaction was stopped by adding 3 ml of ice-cold stop solution containing 100 mM mannitol, 150 mM NaCl and 20 mM HEPES-Tris (pH 7.5), and immediately filtered through a Millipore filter (DAWP, 0.65 μm, 2.5 cm diameter), followed by washing two times with 3 ml of stop solution. Total radioactivity on the washed filters was determined with a liquid scintillation analyzer. S-8921, dissolved in dimethyl sulfoxide, was added to the vesicles and the final concentrations of S-8921 and dimethyl sulfoxide were 0 to 100 μM and 1.6%, respectively.
Analytical methods.
Samples (0.1 ml) of plasma and lymph were added to 10 ml of scintillation fluid (PICO-FLUOR 40, Packard Instrument, Downers Grove, II). Part of the blood, liver homogenate and feces was combusted by use of a sample oxidizer (Tri-Carb, Model 306, Packard Instrument). Radioactivity was measured with a liquid scintillation counter (Tri-carb, Model 2000CA, Packard Instrument). For measurement of [14C]cholesterol and [14C]cholesteryl ester, the plasma and lymph were extracted twice with 6 ml of hexane after addition of 2 ml of ethanol. Extract was evaporated after the total radioactivity had been measured by scintillation counting, and analyzed by TLC [Merck silica gel 60 plate, petroleum ether-acetone (4:1 v/v)]. Protein was determined by the method of Lowry et al. (1951).
Statistical analysis.
Results are expressed as mean ± S.D. Comparisons between groups were made by the Student’st test and Dunnett’s multiple comparison tests. The area under the plasma radioactivity concentration-time curve was obtained by the trapezoidal method.
Results
Hypocholesterolemic effect of S-8921.
As shown in figure2, plasma total and VLDL + LDL cholesterol levels were markedly increased in rats fed a diet containing cholesterol and CA for 7 days, whereas the HDL cholesterol level was slightly decreased but the HDL cholesterol level was very low compared with the VLDL + LDL cholesterol level. Thus, the increase of total cholesterol level by feeding of cholesterol-CA was reflected by the VLDL + LDL cholesterol level. In rats with diet-induced hypercholesterolemia, chronic administration of S-8921 reduced the plasma total cholesterol level, mainly the VLDL + LDL cholesterol level, dose-dependently in the daily dose range of 0.1 to 10 mg/kg. S-8921 suppressed the diet-induced increment of total cholesterol by 21%, 58% and 70% after 3-day treatment at daily doses of 0.1, 1 and 10 mg/kg, respectively. The suppression of total and VLDL + LDL cholesterol levels by S-8921 was already almost maximum after 3 days of treatment, and decreased cholesterol levels were maintained at similar levels throughout the drug administration period. S-8921, given orally at 10 mg/kg, significantly increased HDL cholesterol level after 3 days of treatment .
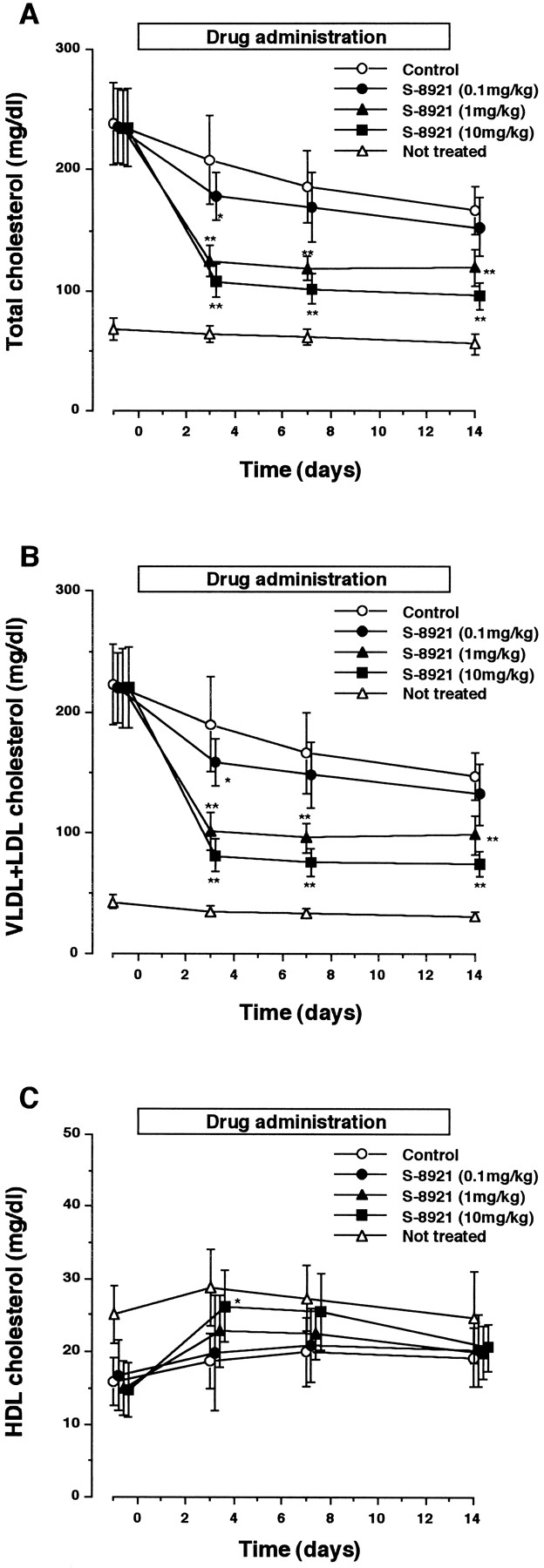
Effect of S-8921 on plasma total cholesterol (A), VLDL + LDL cholesterol (B) and HDL cholesterol (C) in cholesterol-fed rats. Rats were fed a diet containing 1% cholesterol and 0.5% sodium cholate for 7 days, and then S-8921 or vehicle was administered orally for 14 days with cholesterol-cholate feeding. Plasma cholesterol level in normal-diet fed and nontreated rats was also determined. Each value represents the mean ± S.D. of eight rats. Statistically significant difference from control group at *P < .05 and **P < .01 by Dunnett’s multiple comparison test.
Effect of S-8921 on cholesterol absorption and elimination (dual-isotope plasma ratio method).
Figure3 gives the time courses of the plasma radioactivity concentration after coadministration of [14C]cholesterol-labeled chylomicrons given intravenously and of [3H]cholesterol in emulsion given orally after the oral administration of S-8921 to rats. A large fraction (70–98%) of the 14C and3H in the plasma was caused by total cholesterol, and 70 to 86% of the total cholesterol was cholesteryl ester. The plasma 14C concentration after the i.v. injection decreased rapidly until 4 hr after dosing, and after that decreased very slowly. In S-8921-treated rats, the plasma14C concentration decreased significantly compared with control rats on and after 2 days. Also, the14C in the liver at 6 days after the i. v. injection of [14C]cholesterol to control rats and the rats treated with 1 mg/kg and 10 mg/kg of S-8921 were 6.2 ± 2.5%, 2.3 ± 0.4% and 0.8 ± 0.2% of dose, respectively. These values in the rats treated with 1 mg/kg and 10 mg/kg of S-8921 were significantly different from control rats at P < .05 and P < .01, respectively. These results suggest that S-8921 enhanced the elimination of cholesterol from the body. The extents of absorption of cholesterol obtained from the area under the plasma radioactivity concentration-time curves after the coadministration of [14C]cholesterol and [3H]cholesterol to control rats and rats treated with 1 mg/kg and 10 mg/kg of S-8921 were 58.8 ± 6.4%, 32.4 ± 1.4% and 25.1 ± 5.7%, respectively (table1). S-8921 inhibited the absorption of cholesterol from the intestine. Thus, a significant lowering of the plasma radioactivity concentration observed after the oral administration of [3H]cholesterol to S-8921-treated rats appeared to be induced by both actions of inhibiting cholesterol absorption and enhancing of cholesterol elimination.

Effects of S-8921 on time course of plasma radioactivity concentration after coadministration of [14C]cholesterol-labeled chylomicrons, intravenously (A) and of [3H]cholesterol in emulsion, orally (B). S-8921 was administered orally to rats once a day for 9 successive days. On the third day, the isotopes were given 2 hr after administration of S-8921. Each value represents the mean ± S.D. of three to five rats. Statistically significant different from control group at *P < .05 and **P < .01 by Dunnett’s multiple comparison test.
Effect of S-8921 on cholesterol absorption (in situloop method).
Figure 4 shows the effect of S-8921 on the lymphatic absorption of cholesterol after the administration of [14C]cholesterol into the jejunal loops as a mixed micellar solution. When [14C]cholesterol was administered into the jejunal loops of control rats, 30.8 ± 8.9% of the dose was recovered in the thoracic duct lymph at 6 hr. Most (about 90%) of the radioactivity that appeared in the lymph was because of cholesteryl ester. When [14C]cholesterol was administered into the jejunal loops of S-8921-treated rats, the extent of absorption of cholesterol at 2 hr after dosing was low, but those at 4 hr and 6 hr after dosing were not significantly different from those of control rats. The direct inhibitory effect on cholesterol absorption of S-8921 was very weak.

Effect of S-8921 on lymphatic absorption of cholesterol. S-8921 was administered to the jejunal loops of rats at a dose of 10 μmol as an aqueous suspension. After 2 hr, [14C]cholesterol (2 μmol) was administered to the same loop as a micellar solution. Each value represents the mean ± S.D. of four to eight rats. Statistically significant difference from control group at *P < .05 by Student’s t test.
Effect of S-8921 on bile acid excretion (in vivo).
When [14C]TCA was administered orally to control rats (n = 3), 35.1 ± 2.7% of the dose was recovered in the feces at 2 days after dosing, whereas 78.6 ± 4.2% of the dose was recovered in S-8921-treated rats (n = 3). Thus, S-8921 significantly (P < .01) increased the fecal excretion of orally administered [14C]TCA.
Effect of S-8921 on bile acid absorption (in situ loop method).
Figure 5 shows the cumulative extent of absorption of the radioactivity in the mesenteric venous blood after the administration of [14C]TCA or [14C]GCA into the rat ileal loops. When [14C]TCA was administered into the ileal loops of control rats, 75.7 ± 12.4% of the dose was absorbed in the mesenteric blood at 30 min after dosing, and 46.5 ± 11.7% of the dose was absorbed in case of [14C]GCA, whereas the extents of absorption for both bile acids were very low in rats to which S-8921 had been given in the ileal loops. On the other hand, when [14C]TCA was administered into the ileal loops of rats to which S-8921 had been given intravenously at the same dose as that of ileal administration, 78.5 ± 11.4% of the dose was absorbed at 30 min after dosing. Thus, S-8921 given orally may directly act on the intestinal lumen because it had no significant effect on TCA absorption when given intravenously. Figure6 shows the cumulative extent of absorption of the radioactivity in the mesenteric blood after administration of [14C]CA into the ileal and jejunal loops of rats. When [14C]CA was administered into the ileal loops of control rats, 53.5 ± 13.2% of the dose was absorbed at 30 min after dosing, whereas only 9.7 ± 4.4% of the dose was absorbed in S-8921-treated rats. In contrast, when [14C]CA was administered into the jejunal loops of control and S-8921-treated rats, the extents of absorption were 21.2 ± 10.1% and 21.2 ± 2.5% at 30 min after dosing, respectively. The extents of absorption from the jejunum of [14C]CA were not significantly different between the two groups. Thus, when S-8921 was given in situin the intestinal loops, it inhibited the active absorption of bile acids from the rat ileum but not the passive absorption of CA from the rat jejunum.

Effects of S-8921 on portal absorption of TCA (A) and GCA (B) from the terminal ileum. S-8921 was administered to the ileal loops of rats at a dose of 0.5 μmol as an aqueous suspension. After 1 hr, [14C]bile acid aqueous solution (5 μmol) was administered to the same loop. Each value represents the mean ± S.D. of three rats. Statistically significant difference from control group at *P < .05 and **P < .01 by Student’st test.

Effects of S-8921 on portal absorption of CA from the terminal ileum (A) and the jejunum (B). S-8921 was administered to the intestinal loop of rats at a dose of 0.5 μmol as an aqueous suspension. After 1 hr, [14C]CA aqueous solution (5 μmol) was administered to the same loop. Each value represents the mean ± S.D. of three rats. Statistically significant difference from control group at **P < .01 by Student’s ttest.
Effect of S-8921 on bile acid transport (in vitro).
The time courses for uptake of TCA,d-glucose and l-alanine by brush-border membrane vesicles isolated from the jejunum and ileum of the rat small intestine in the presence of a Na+ gradient are shown in figure 7. In the brush-border membrane vesicles isolated from the ileum, the uptake of TCA was transient, reaching a maximum accumulation of about 0.6 nmol/mg of protein at 2 min and then decreasing to about 0.2 nmol/mg of protein during the next 10 to 30 min (overshoot phenomenon). In contrast, in the brush-border membrane vesicles isolated from the rat jejunum, no overshoot uptake of TCA was observed. The Na+-dependent overshoot uptake ofd-glucose was observed in the brush-border membrane vesicles isolated from the rat jejunum, and the d-glucose uptake for 15 sec was 4 times greater than that observed at 2 min. Also, the Na+-dependent overshoot uptake ofl-alanine was observed in the brush-border membrane vesicles isolated from the rat jejunum and ileum. These overshoot uptakes of TCA, d-glucose and l-alanine were observed only in the presence of Na+ gradient, and these uptakes in the absence of Na+ were very low. As shown in table 2, S-8921 decreased the Na+-dependent uptake of TCA by the rat ileal brush-border membrane vesicles by 25 to 98% at concentrations of 1 to 100 μM. S-8921 inhibited the uptake of TCA by the rat ileal brush-border membrane vesicles in a concentration-dependent manner with IC50 of 2.1 μM. In contrast, S-8921 did not inhibit the Na+-dependent d-glucose uptake by the rat jejunal brush-border membrane vesicles and the Na+-dependent l-alanine uptake by the rat jejunal and ileal brush-border membrane vesicles at the concentration of 100 μM.

Uptake of TCA (A), d-glucose (B) andl-alanine (C) by brush-border membrane vesicles from the rat jejunum and ileum. Uptake was initiated at zero time by addition of a medium (70 μl) containing 100 mM mannitol, 150 mM NaCl, 0.05 mM [14C]TCA and 20 mM HEPES/Tris buffer (pH 7.5), or containing 100 mM mannitol, 0.1 mM MgSO4, 100 mM NaSCN, 1 mM HEPES/Tris buffer (pH 7.5), 22.9 μMd-[14C]glucose or 0.008 μMl-[14C]alanine to vesicles (50 μl).
Effects of S-8921 on uptake of TCA, d-glucose andl-alanine by the rat ileal or jejunal brush-border membrane vesicles
Discussion
The present study examined the mechanism of the hypocholesterolemic action of S-8921, a new compound, in rats. In rats with diet-induced hypercholesterolemia, chronic oral administration of S-8921 lowered plasma total cholesterol, mainly the VLDL + LDL cholesterol level. To determine whether this suppression of plasma cholesterol by S-8921 is caused by inhibition of cholesterol absorption or enhancement of cholesterol elimination, we investigated the effect of S-8921 on cholesterol absorption and disposition by the dual-isotope plasma ratio method (Zilversmit, 1972; Zilversmit and Hughes, 1974). The extents of absorption of cholesterol obtained from the area under the plasma radioactivity concentration-time curves after coadministration of [14C]cholesterol-labeled chylomicrons intravenously and of [3H]cholesterol orally decreased with S-8921 treatment. Also, S-8921 enhanced the elimination of cholesterol from the body. These results point to a significant lowering of the plasma radioactivity concentration after oral administration of [3H]cholesterol to S-8921-treated rats. Thus, the suppression of plasma total and VLDL + LDL cholesterol levels by S-8921 are caused by both inhibition of cholesterol absorption and enhancement of cholesterol elimination. On the other hand, when [14C]cholesterol was administered into the jejunal loops of control and S-8921-treated rats, the lymphatic absorption of cholesterol was not significantly different between the two groups. This finding suggests that the inhibitory effect of S-8921 on cholesterol absorption is not simply the result of direct inhibition of cholesterol absorption in the jejunum. Absorption of dietary cholesterol depends on several factors of which the bile acid concentration in the intestine is of major importance. Also, cholesterol excretion from the body occurs almost exclusivelyvia the hepatobiliary system as the sterol itself or its major metabolites, the bile acids (Packard and Shepherd, 1982). Interruption of the enterohepatic circulation of bile acids by bile acid sequestrants (Glueck et al., 1972) or partial ileal bypass surgery (Buchwald et al., 1990) can cause a significant decrease of plasma LDL cholesterol concentrations. S-8921 increased the fecal excretion of orally administered [14C]TCA. The hypocholesterolemic action of S-8921 may be attributed to interruption of the enterohepatic circulation of bile acids by this drug. Therefore, the effect of S-8921 on bile acid absorption from the intestine was examined. When conjugated bile acids such as TCA or GCA or the free bile acid CA were administered into the ileal loops of rats, their extents of absorption in the mesenteric blood were effectively lowered by pretreatment with S-8921. In contrast, when CA was administered into the jejunal loops of control and S-8921-treated rats, no significant difference of the extents of absorption between these two groups was observed. The conjugated bile acids such as TCA and GCA are mostly absorbed by Na+-dependent active transport in the terminal ileum, whereas free bile acids such as CA are absorbed to a much greater extent by passive diffusion throughout the entire intestine (Holt, 1966; Lack and Weiner, 1966; Lack et al., 1970; Krag and Phillips, 1974). Thus, S-8921 may selectively inhibit the active absorption of bile acids from the rat ileum but not inhibit the passive absorption of bile acids.
Bile acids are synthesized from cholesterol in the liver and secreted into the small intestine. More than 90% of the bile acids are reabsorbed either by passive diffusion along the entire intestine or by active transport at the terminal ileum (Holt, 1966; Lack et al., 1970). Because the bile acid pool is tightly regulated, if the enterohepatic circulation of bile acids is interrupted in some manner, their biosynthesis from cholesterol would increase to compensate for the loss in the liver. Partial depletion of hepatic cholesterol by the enhancement of cholesterol catabolism causes increased uptake of plasma LDL cholesterol via liver LDL receptor (Packard and Shepherd, 1982). Furthermore, decrease of bile acid concentration in bile leads to less cholesterol absorption in the small intestine. The anion exchange resins cholestylramine and colestipol are the only drugs in clinical use which have this mechanism of action (Ast and Frishman, 1990), and their combined use with other hypocholesterolemic agents, such as HMG CoA reductase inhibitors, is more effective (Vega and Grundy, 1987; Leren et al., 1988). However, the drawbacks of bile acid sequestrants include the discomfort of intake owing to their bulkiness and unpalatable texture and their having gastrointestinal side effects, which results in poor patient compliance (Hunninghake, 1986). Therefore, it would be desirable to develop a hypocholesterolemic agent that can interrupt the reabsorption of bile acids by the ileal bile acid active transport system at a low dose. Recently, inhibitors of the ileal bile acid active transport system have been found (Fears et al., 1990; Wess et al., 1994; Root et al., 1995), and several inhibitors decrease plasma LDL cholesterol concentrations in rats (Fears et al., 1990; Lewis et al., 1995).
The present study has shown that S-8921 is also a specific inhibitor of the ileal bile acid transport system. Studies with isolated membrane vesicles have indicated that a transporter associated with the ileal brush-border membrane is responsible for the binding and active cotransport of bile acids with sodium ions (Wilson, 1981; Krameret al., 1993). Recently, cDNA cloning of ileal Na+-dependent bile acid transporter protein was performed for hamsters (Wong et al., 1994), humans (Wonget al., 1995) and rats (Schneider et al., 1995). To locate the site of inhibition of S-8921 within the active transport system for bile acids, we investigated the effect of S-8921 on TCA uptake by ileal brush-border membrane vesicles prepared from rat terminal ileum. We found that S-8921 inhibited this Na+-dependent uptake of TCA in a concentration-dependent manner. As with the transport of bile acids,d-glucose and amino acids are generally considered to be transported by Na+-dependent intestinal transport systems (Hopfer et al., 1976; Crane, 1977; Kessler et al., 1978). However, S-8921 did not inhibit the Na+-dependent d-glucose uptake by the rat jejunal brush-border membrane vesicles and the Na+-dependent l-alanine uptake by the rat jejunal and ileal brush-border membrane vesicles. These results suggest that S-8921 is a potent selective inhibitor of the Na+-dependent bile acid transporter in the ileal mucosal cell brush-border membrane, and this inhibition is the mechanism by which this drug suppresses the reabsorption of bile acids in the ileum. We have observed that S-8921 can inhibit the uptake of TCA in the COS7 cell line which constitutively expresses hamster ileal Na+-dependent bile acid transporter (Haraet al., 1997).
In conclusion, S-8921 is a potent hypocholesterolemic agent for rats with diet- induced hypercholesterolemia. It may act by inhibiting cholesterol absorption in the intestine and by enhancing cholesterol elimination from the body by interrupting the reabsorption of bile acids based on selective inhibition of the Na+-dependent bile acid transport in the mucosal cell brush-border membrane. Specific inhibitors of ileal bile acid transport system, such as S-8921, show promise as a new class of hypocholesterolemic drugs.
Acknowledgment
The authors thank Drs. Seijiro Hara and Sachio Mori for their many helpful comments and suggestions on the preparation of the manuscript.
Footnotes
-
Send reprint requests to: Dr. Teruhisa Ichihashi, Formulation R & D Laboratories, Shionogi & Co., Ltd., Fukushima-ku, Osaka 553, Japan.
- Abbreviations:
- TCA
- taurocholic acid
- GCA
- glycocholic acid
- CA
- cholic acid
- VLDL
- very low-density lipoprotein
- LDL
- low-density lipoprotein
- HDL
- high-density lipoprotein
- HMG CoA
- 3-hydroxy-3-methylglutaryl coenzyme A
- HEPES
- N-(2-hydroxyethyl)-piperazine-N′-2-ethanesulfonic acid
- TLC
- thin-layer chromatography
- Received March 31, 1997.
- Accepted September 15, 1997.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}